Функции нерва полярный закон возбуждения нерва
Содержание статьи
4. Законы проведения возбуждения по нервам.
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну.Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках — от рецептора к клетке, в эфферентных — от клетки к рабочему органу.
Закон анатомической ифизиологической целостности нервного волокна.Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированногопроведения возбуждения по нервномуволокну.В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
Возбуждение (потенциал действия) распространяется по нервному волокну без затухания.
Периферический нерв практически неутомляем.
Механизм проведения возбуждения по нерву.
Возбуждение (потенциал действия — ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным.
Скорость распространения возбуждения пропорциональна толщине волокна и обратно пропорциональна сопротивлению среды. Проведение возбуждения зависит от соотношения амплитуды ПД и величины порогового потенциала. Этот показатель называется гарантийный фактор(ГФ) и равен 5 — 7, т.е. ПД должен быть выше порогового потенциала в 5- 7 раз. Если ГФ = 1 проведение ненадёжно, если ГФ < 1 проведения нет. Протяженность возбуждённого участка нерва L является произведение времени (длительности) ПД и скорости распространения ПД. Например, в гигантском аксоне кальмара L= 1 мс25 мм/мс = 25 мм.
Наличие у миелиновых волоконоболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки — перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Вмиелинизированном волокне токи проводятся только в зонах, не покрытых миелином, — перехватах Ранвье, в этих участках и генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 — 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. Распространение ПД в миэлинизированных нервных волокнах осуществляетсясальтаторно- скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Скорость такого способа проведения возбуждения значительно выше, и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов, благодаря чему уменьшается нагрузка на ионный насос.

Схема распространения возбуждения в безмиелиновых и миелиновых нервных волокнах.
Соседние файлы в предмете Нормальная физиология
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
9. Законы раздражения. Полярный закон. Физиологический электротон. Като-дическая депрессия.
Полярный закон раздражения (закон Пфлюгера)

При замыкании цепи постоянного электрического тока возбуждение возникает только под катодом, а при размыкании — только под анодом
Доказать правильность закона можно при помощи опыта, в котором меняется расположение катода и анода на поврежденном и неповрежденном участках нерва, иннервирующего мышцу
Прохождение постоянного электрического тока через мембрану вызывает изменение мембранного потенциала покоя.
Так, при замыкании цепи возле катода скапливается его «-» заряд, который уменьшает «+» заряд наружной поверхности мембраны.
Разность потенциалов (между наружной и внутренней поверхностями мембраны) уменьшается, и мембранный потенциал изменяется в сторону уровня критической деполяризации, т.е. формируется деполяризация(возбудимость повышается).
Достижение критического уровня приводит к возникновению пикового потенциала (потенциала действия)
При замыкании цепи возле анода скапливается его «+» заряд.
Он увеличивает «+» заряд мембраны и величину мембранного потенциала
Мембранный потенциал удаляется от критического уровня, превышает значение потенциала покоя и формируетгиперполяризацию (возбудимость уменьшается)
При размыкании цепи, прекращение поступления дополнительного «+» заряда от анода приводит к уменьшению(восстановлению) заряда наружной поверхности мембраны.
Мембранный потенциал, уменьшаясь, приближается к критическому уровню
Формируется деполяризация (возбудимость увеличивается).
После достижения критического значения развивается пиковый потенциал
(Поскольку размыкание происходит после замыкания, а, следовательно, на фоне гиперполяризации и пониженной возбудимости, то для возникновения ПД необходим раздражитель, превышающий по силе пороговый — это анодно-размыкательный эффект)
При размыкании возле катода прекращается накопление его «-» заряда
Заряд наружной поверхности мембраны увеличивается (восстанавливается), мембранный потенциал, увеличиваясь, удаляется от критического уровня и возбуждение не возникает
Закон физиологического электротона
Действие постоянного электрического тока на ткань сопровождается изменением ее возбудимости
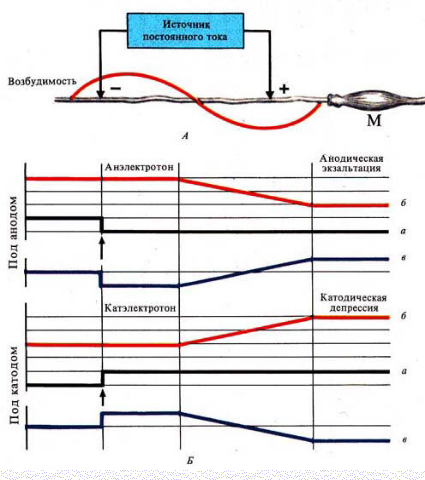
Различают 3 вида физиологического электротона или изменения возбудимости:
Катэлектротон — изменение возбудимости под катодом.
В момент замыкания под катодом формируется деполяризация и возбудимость повышается.
По мере удаления от катода количество его «-» зарядов, а следовательно и выраженность деполяризацииуменьшается.
В результате возбудимость уменьшается, но она остается выше, чем в состоянии покоя
Анэлектротон — изменение возбудимости под анодом.
В момент замыкания под анодом формируется гиперполяризация и возбудимость снижается.
По мере удаления от анода количество его «+» зарядов, а, следовательно, и гиперполяризация уменьшается.
В результате возбудимость увеличивается, но она остается ниже, чем в состоянии покоя
Периэлектротон — обратное изменение возбудимости вне электротонических областей.
В зоне прекращения действия катода возбудимость снижается.
В зоне прекращения влияний анода она, наоборот, увеличивается
При длительном прохождении постоянного тока через ткань происходит извращение измененной возбудимости
При длительном замыкании под катодом происходит:
·инактивация Na проницаемости
·повышение уровня критической деполяризации
·увеличение порога деполяризации
·снижение вначале повышенной возбудимости
Это явление получило название катодическая депрессия
При длительном замыкании под анодом происходит:
·снижение К проницаемости
·уровень критической деполяризации снижается
·порог деполяризации уменьшается
·повышается вначале сниженная возбудимость
Это явление получило название анодическая экзальтация
Этот закон необходимо учитывать в медицинской практике
С одной стороны, он может быть использован, если требуется заблокировать проведение возбуждения по нервной или мышечной ткани (катодическая депрессия) или повысить возбудимость (анодическая экзальтация)
С другой стороны, необходимо помнить о возможности привыкания к длительному действию раздражителей, в частности, фармацевтических препаратов, влияющих на возбудимость мембраны.
ФИЗИОЛОГИЯ МЫШЦ И НЕРВОВ
Структура миофибрилл и ее изменения при сокращении.Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнахмиофибриллы-разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны,т. е. обладают двойным лучепреломлением. В обыкновенном свете они ^выглядят темными, а. в поляризованномг- прозрачными в продольном направлении й непрозрачными в поперечном. Другие участки в Обыкновенной свете выглядят светлыми — они изотропны,т. е. не ; обладают двойным’ лучепреломлением (рис. 34, а) . Анизотропные участки обозначают буквой А,изотропные — буквой I.В середине диска Аразличается светлая полоска Н, посередине диска / — темная «полоска -представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по,отношению друг к другу.
Современные представления о структуре миофибриллярного аппарата основываются на исследованиях структуры мышечного волокна-при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл,представляющих собой удлиненные полимери-, зованные молекулы белков миозина и актина; Миозиновые протофибриллы, или, как, их принято обозначать, нити, вдвое толще актиновых. Их диаметр,примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, ^что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б). Благодаря этому диски / состоят только из актиновых нитей, а диски А — из нитей миозина, а, возможно, еще и другого белка.

Рис. 34. Структура миофибриллы (схема), Показаны диски А и 1-полоски Zи Н(а). Взаимное расположение толстых (миозиновых) и тонких ; (актиновых) нитей в расслабленной (б) й сокращённой (в) миофибрилле.
Светлая полоска Н представляет собой узкую зону, свободную от актиновых нйтей. Мембрана Z,проходя через середину диска /, скрепляет,между собой эти Нити. Важным компонентом ультрамикроскопической-структуры миофибрилл являются также многочисленные поперечные мостики, .соединяющие между собой миозиновые и актиновые нити. При сокращений мышечного волокна указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего диски / укорачиваются, а диски/1 сохраняют свой размер. Почти исчезает светлая полоска //, так как актиновые нити при сокращении сближаются друг с другом своими концами (рис. 34, в). Причиной «скольжения» является химическое взаимодействие между актином и миозином в присутствии ионов Са2+и АТФ. Наблюдается своегородачхимическое «зубчатое колесо», как 6^,1 протягивающее одну Группу нитей по другой. Роль «зубчиков» в этом процессе приписывают поперечным мостикам, обеспечивающим взаимодействие активных центров белков миозиновых и актиновых нитей.
Молекула актина
Это глобулярный белок, состоящий из одного полипептида, который полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга (рис. 4-4 А). Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся мышечном волокне взаимодействие между актином и миозином предотвращают два белка — тропонин и тропомиозин (рис. 4-4 Б).

Тропомиозин — стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин — гетеротримерный белок. Он состоит из тропонина Т (отвечает за связывание с одиночной молекулой тропомиозина), тропонина С (связывает ион Са2+) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
Миозин
Это единое название большой семьи протеинов, имеющих определенные отличия в клетках разных тканей. Миозин присутствует у всех эукариотов. Около 60 лет назад было известно два типа миозина, которые сейчас называют миозин I и миозин II. Миозин II был первым из числа открытых миозинов, и именно он принимает участие в мышечном сокращении. Позднее были открыты миозин I и миозин V (рис. 4-4 В). В последнее время показано, что миозин II участвует в мышечном сокращении, тогда как миозин I и миозин V вовлечены в работу подмембранного (кортикального) цитоскелета. В настоящее время идентифицировано более 10 классов миозина. На рисунке 4-4 Г показано два варианта схемы строения миозина, который состоит из головки, шейки и хвоста. Молекула миозина состоит из двух больших полипептидов (тяжелых цепей) и четырех меньших (легких цепей). Эти полипептиды составляют молекулу с двумя глобулярными «головками», которые содержат оба вида цепей, и длинным стержнем («хвостом») из двух переплетенных тяжелых цепей. Хвост каждой молекулы миозина располагается вдоль оси толстого филамента, а две глобулярные головки выступают по бокам, их иначе называют поперечными мостиками. На каждой глобулярной головке находятся по два участка связывания: для актина и для АТФ. Участки связывания АТФ обладают также свойствами фермента АТФазы, гидролизующей связанную молекулу АТФ.
Источник
ГЛАВА 5. ФИЗИОЛОГИЯ НЕЙРОНОВ
Нервная система состоит из двух типов клеток — нервных (нейроны) и глиальных (нейроглия). К нейроглии относятся астроциты, микроглия, миелинобразующие клетки (олигодендроциты ЦНС и шванновские клетки периферических нервов). По отношению к нейронам глиоциты выполняют трофическую, опорную и изолирующую (электрический изолятор) функции. Нейрон — основная структурно-функциональная единица нервной системы. Количество нейронов в мозге человека превышает 100х109. Основная функция нейронов — генерация, передача и интеграция нервных импульсов.
Нейроны
В нервной клетке различают тело (перикарион) и отростки — дендриты и аксон (рис. 5-1). Как правило, нейрон имеет несколько дендритов и один аксон. Отростки нейрона и поверхность перикариона участвуют в образовании синапсов. Нейроны — возбудимые клетки — осуществляют передачу электрических сигналов в пределах самой нервной клетки (а между нейронами при помощи нейромедиаторов в синапсах) и этим обеспечивают способность мозга к переработке информации. Сома нейрона синтезирует белки и различные клеточные материалы и распределяет их по дендритам и аксону.
Транспорт веществ в нейроне и его отростках. Транспортные механизмы способны перемещать клеточные компоненты в антероградном направлении в сторону от сомы нейрона или в ретроградном направлении в сторону сомы нейрона. Кинезин и микротубулоассоциированный белок ответственны за антероградный аксональный транспорт органелл и везикул; динеин и другой микротубулоассоциированный белок отвечают за ретроградный транспорт. Антеро-
Рис. 5-1. Многоотростчатый нейрон [11]. Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них — аксон, все другие — дендриты. Справа и сверху вниз: функциональные зоны нервной клетки — рецептивная (дендритная), аксон (область проведения ПД), концевых разветвлений аксона (пресинаптическая).
градный транспорт подразделяется на быстрый и медленный. Скорость медленного аксоплазматического транспорта 1-2 мм в день. Скорость быстрого аксоплазматического транспорта около 400 мм в день. Быстрый транспорт используется для переноса органелл, везикул и мембранных гликопротеинов в терминаль аксона. Медленный транспорт используется для переноса структурных белков, таких как актин, нейрофиламенты и микротубулы. Ретроградный аксональный транспорт обеспечивает доставку к нейрону трофи- ческих факторов, например фактора роста нервов.
Классификации нейронов
Существует множество классификаций нервных клеток, так как нейроны различаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в нейронных сетях и по множеству других признаков. В зависимости от класса выполняемой функции выделяют афферентные (чувствительные, сенсорные), эфферентные (двига-
тельные, моторные) и вставочные нервные клетки (ассоциативные нейроны, или интернейроны).
Исходя из характера электрогенеза в различных зонах нервных клеток (рецептивная и интегративная зоны, начальный сегмент ак- сона, аксон, терминали аксона) предложена универсальная классификация (см. рис. 5-1). Эта классификация учитывает направление (вход и выход) сигналов в пределах одной нервной клетки, но находящейся в составе нейронных сетей или цепочек, а также характер мембранного электрогенеза (изменения МП в рецептив- ной и пресинаптической зонах и передача ПД по аксону).
Мембранный потенциал покоя
Мембранный потенциал (МП) покоя — разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя (при отсутствии раздражителей) — рассмотрен в главе 2 (раздел «Мембранный электрогенез»). МП покоя нервной клетки составляет -70 мВ, и его величина, устанавливаемая безворотными ионными каналами, довольно стабильна. Изменения величины МП (рис. 5-2) описываются специальными терминами:
• деполяризация — уменьшение величины МП;
• реполяризация — возвращение величины МП к исходному уровню;
• гиперполяризация — увеличение МП по сравнению с исходным уровнем.
Все изменения МП в дендритной зоне нервной клетки, формируемые при участии воротных каналов, которые управляются лигандами, известны как локальные ответы, затухающие на протяжении нескольких микрометров от места возникновения. В этой зоне постоянно возникают десятки и сотни локальных ответов (постсинаптические потенциалы), суммирование которых в виде изменений МП достигает области начала аксона (аксонного хол- мика). Помимо локальных ответов, возбудимые мембраны нервных клеток в области аксона за счет быстрой активации множества потенциалзависимых натриевых каналов могут генерировать потенциалы действия (ПД, спайки, нервные импульсы, см. рис. 5-2). Существенное значение при генерации ПД имеет представление о пороге раздражения.
Рис. 5-2. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке -80 мВ — исходный уровень МП.
Локальные ответы, порог раздражения и потенциал действия
Нервная клетка на раздражающие (в том числе электрические) стимулы отвечает изменениями МП: либо реагирует локальными (подпороговыми) реакциями, либо генерирует ПД.
• Локальные ответы. Стимулы, величина которых меньше опре- делённого порога, вызывают локальные ответы, затухающие вблизи места раздражения. Эти локальные ответы могут быть как деполяризующими (возбуждающие) мембрану, так и гиперполяризующими (тормозные).
• Потенциал действия. Деполяризующие стимулы, достигшие порогового значения, вызывают развитие возбуждения в виде ПД. ПД распространяется по мембране нейрона без затухания (без декремента).
Законы раздражения нервной клетки
Генерация ПД подчиняется законам «всё или ничего», «сила- время» и аккомодации.
• Закон «всё или ничего». При достижении пороговой силы раздражающего стимула дальнейшее увеличение его интенсивности или продолжительности раздражения не изменяет ха-
рактеристик ПД. Такая форма реакции нервной клетки на раздражение получила название «всё или ничего» (либо ПД возникает, либо нет).
• Закон силы-времени. Эффект раздражителя на нервную клетку зависит не только от силы раздражителя, но и от времени, в течение которого он действует; так, чем больше сила тока, тем меньше времени он должен действовать, чтобы возник процесс возбуждения, и наоборот. Соотношение силы и длительности действия раздражителя может быть выражено в виде гиперболической кривой (рис. 5-3).
• Закон аккомодации. Раздражающее действие тока зависит не только от амплитуды электрического сигнала, но и от скорости (крутизны) его нарастания во времени: чем быстрее это нарастание, тем сильнее выражено раздражающее действие тока. При действии медленно нарастающей силы раздражителя генерации ПД не происходит, так как возбудимая клетка
в
Рис. 5-3. Кривая зависимости силы раздражения возбудимой ткани от продолжительности действия раздражителя «сила-длительность» [21]. 1 — реобаза (минимальная сила раздражителя, способная вызвать ПД); 2 — удвоенная реобаза; 3 — кривая «сила-длительность»; а — полезное время (минимальное время, в течение которого ток, равный одной реобазе, вызывает генерацию ПД); б — хронаксия (минимальное время, в течение которого ток, равный двум реобазам, вызывает ПД).
«приспосабливается» к действию этого раздражителя, что и получило название аккомодации.
Потенциал действия
Потенциал действия (ПД) — быстрое колебание МП — самораспространяющийся процесс, связанный с изменениями ионной проводимости мембраны, которые вызваны функционированием потенциалзависимых натриевых каналов. ПД распространяется без затухания, т.е. практически без уменьшения амплитуды. Проведение ПД по мембране можно сравнить с поджиганием пороховой дорожки: вспыхнувший порох немедленно воспламеняет впереди лежащие частицы, и пламя движется вперёд до конца дорожки.
Временной ход потенциала действия. Продолжительность ПД нервной клетки измеряется единицами миллисекунд (мс). ПД, зарегистрированные двумя электродами, один из которых находится внутри клетки, а другой — в окружающем растворе, представлены на рис. 5-2, 5-4.
Рис. 5-4. Потенциал действия и ионные токи нервной клетки [5]. По оси
ординат отложены значения МП (мВ), по оси абсцисс — время (мс).
Между моментом, когда наносится раздражение, и первым проявлением ПД существует задержка — латентный период. Латент- ный период соответствует времени, когда ПД движется по мембране нервной клетки от места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны — локальный ответ. При достижении критического уровня деполяризации, который в среднем составляет -55 мВ, начинается фаза деполяризации. В этой фазе уровень МП падает до нуля и даже приобретает положительное значение (овершут), а затем возвращается к исходному уровню (фаза реполяризации). Фазы деполяризации, овершута и реполяризации образуют спайк (пик) ПД. Длительность спайка составляет 1-2 мс. После спайка наблюдается замедление скорости спада потенциала — фаза следовой деполяризации. После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации. Эти следовые потенциалы могут длиться десятки и сотни миллисекунд.
Ионные механизмы потенциала действия. В течении ПД происходят изменения МП, обусловленные ионными механизмами. На рис. 5-4 представлены суммарные ионные токи, протекающие через мембрану нервной клетки в ходе ПД.
• Локальный потенциал. Раздражение клетки обусловливает открытие части потенциалзависимыхNa+-каналов и появление локального (нераспространяющегося) потенциала.
• Фаза деполяризации. При достижении критического уровня деполяризации мембраны (соответствует порогу активации потенциалзависимых Na+-каналов) начинается лавинообразный процесс открытия множества Na+-каналов. В фазе деполяризации происходит массивный вход в клетку ионов Na+ по концентрационному и электрохимическому градиенту.
• Овершут. Деполяризация мембраны приводит к реверсии мембранного потенциала (МП становится положительным). В фазе овершута Na+-ток начинает стремительно спадать, что связано с инактивацией потенциалзависимых Na+-каналов (время открытого состояния этих каналов — доли миллисекунды) и дополнительным открытием К+-каналов.
• Фаза реполяризации. Помимо инактивацииNa+-каналов, развитию реполяризации способствует открытие потенциалзависимых К+-каналов. Этот процесс происходит медленнее, чем
открытие Na+-каналов, но К+-каналы остаются открытыми более продолжительное время. Выход К+ наружу способствует полному завершению фазы реполяризации.
• Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов, что способствует выходу ионов К+. Восстановление исходного уровня МП приводит Na+- и К+-каналы в состояние покоя.
Изменения возбудимости во время потенциала действия. В ходе развития ПД изменяется возбудимость мембраны нервной клетки.
• Абсолютно рефрактерный период. Во время фазы деполяризации и частично фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинстваNa+-каналов. Абсолютно рефрактерный период ограничивает частоту возникновения ПД и возможность его проведения в неверном направлении по аксону.
• Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов.
• Лабильность — максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с.
Проведение возбуждения по нервным волокнам
Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских клеток в периферических нервах. Нервные волокна подразделяются на два типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5-5) и в значительной мере зависит от диаметра нервных волокон.
• Безмиелиновые нервные волокна (рис. 5-6, А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется и наружная поверхность мембраны при-
Рис. 5-5. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
обретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны. Такое прове- дение известно как электротоническое, а проведение ПД — своего рода «эстафета», в которой каждый участок мембраны является сначала раздражаемым, а затем раздражающим. ПД возникает за счёт увеличивающейся проводимости через потенциалзависимые Na+-каналы, встроенные в аксолемму с плотностью около 110-120 каналов на 1 мкм2. Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении. Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5-2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения
ПД (см. рис. 5-5).
Миелиновое нервное волокно (рис. 5-6, Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм; это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату ( скачкообразное проведение).
Рис. 5-6. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение); Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье — проводник.
Законы проведения возбуждения
• Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, т.е. проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени.
• Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена:
Φ наличием оболочек, окружающих отдельные нервные волокна и их пучки (в результате образуется барьер, предупреждающий переход возбуждения с волокна на волокно);
Φ сопротивлением межклеточной жидкости (жидкость, находящаяся между волокнами, имеет гораздо меньшее сопротивление току, чем мембрана аксонов; поэтому ток шунтируется по межволоконным пространствам и не доходит до соседних волокон).
Типы нервных волокон и их функции
При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в 1937 г. обнаружили, что ток действия нервного ствола имеет составной характер. На основании полученных данных (диаметр, скорость проведения, функция) разработана классификация (табл. 5-1), в соответствии с которой нервные волокна подразделены на группы A, В и С с дальнейшими градациями (α, β и т.д.).
Таблица 5-1. Классификация нервных волокон по диаметру и скорости проведения [3]
Структуры
Окончание табл. 5-1
Примечание. Классификация Эрлангера-Гассера (латинские буквы) и альтернативные добавления (римские цифры).
Обобщение главы
Нейроны имеют разветвлённую систему дендритов и, как правило, один протяжённый аксон.
Локализация ионных каналов носит избирательный характер: на нейроне каналы, не имеющие ворот, распределены на нейрональной мембране; потенциалзависимые каналы, в основном, располагаются в аксоне и его отростках.
Потенциал действия представляет собой преходящее изменение мембранного потенциала, характеризующееся быстрой деполяризацией и последующей реполяризацией.
Фаза деполяризации — результат быстрой активации потенциалзависимых Na+-каналов; фаза реполяризации — результат инактивации потенциалзависимых Na+-каналов и задержанной активации потенциалзависимых К+-каналов.
Потенциал действия в нейроне возникает тогда, когда аксонный холмик деполяризуется до порогового уровня быстрой активацией большого количества потенциалзависимых натриевых каналов.
Распространение потенциала действия происходит за счёт постоянного его возникновения в соседних участках мембраны вследствие локального тока, возникающего между возбуждённым и невозбуждённым отделами.
Скорость проведения потенциала действия зависит от диаметра аксона и толщины миелиновой оболочки.
Антероградный аксональный транспорт обеспечивает терминали аксона органеллами, синаптическими везикулами, метаболическими факторами и рецепторами.
Ретроградный аксональный транспорт переносит отработанные вещества, захватывает питательные материалы и сигнальные по- средники из области терминали для сомы нейрона и служит механизмом для входа в ЦНС токсинов и микроорганизмов.
Источник